
Cs. Kádár Péter - XXI. századi Diszkónika, 497. Nézz a szemedbe! – második pillantás
A látás sajátosságainak megértéséhez körül kell nézni az agyban.
Az itt végbemenő feldolgozási folyamatok felderítéséhez az egyik leggyakoribb kísérleti eljárás az egy-sejt vizsgálat.
Az agy alapvető funkcionális egységei a neuronok. Neuronnak nevezzük az idegsejt és nyúlványainak együttesét. Az idegsejtet és nyúlványait egybefüggő sejthártya (plazmamembrán) határolja. A neuronok ingerlékeny sejtek, amelyek ingerfelvételre és idegi ingerületek vezetésére specializálódtak. A neuronok hálózatot alkotnak.
A központi idegrendszer és a hozzá kapcsolódó érző ganglionok idegsejtjeinek működésük szerint három fő típusa van. Az érző neuronok az ingerület felvételét és továbbítását végzik. Az interneuronok feladata az ingerület továbbadása, valamint más neuronok közötti kapcsolat fenntartása. A mozgató neuronok pedig az ingerületre adott válaszreakciót valósítják meg.
Az egy-sejt eljárás középpontjában a neuronon keresztülhaladó elektromos áram áll. Ahogy az akciós potenciál (lásd a sorozat egyik legszexisebb, 33. részét) végighalad egy sejten, az áram ki- és befolyik a sejttest és az axon membránfelszínén. Az axon az idegsejt sajátos szerkezetű, leghosszabb nyúlványa.
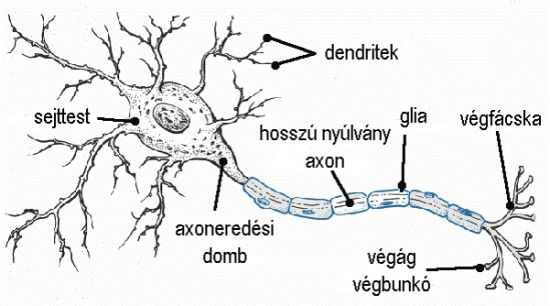
Egy idegsejtnek csak egyetlen axonja van. Az elektromos áramlás mérhető feszültséget hoz létre a sejten belül és kívül egyaránt. Emiatt két alapvető formája különíthető el az egy-sejt vizsgálatoknak. Az intracelluláris egy-sejt vizsgálatok a sejten belül zajlanak, a feszültségváltozást az idő függvényében a sejtmembránon rögzítik az akciós potenciálok jelentkezése során. Az extracelluláris vizsgálat esetében a mikroelektródát a sejtfelszínhez közel helyezik el, így a feszültségváltozásokból csak korlátozott információk nyerhetők. Az egy-sejt vizsgálat kivitelezéséhez alapvetően szükség van mikroelektródákra, erősítőre, mikromanipulátorokra és rögzítő berendezésre. A vizsgálat célja határozza meg a használandó mikroelektróda fajtáját. Az elektródák nagy ellenállása problémát jelent a jel erősítése során. Ha egy alacsony bemenő impedanciájú erősítőhöz kapcsolódna az elektróda, nagy potenciálzuhanás lenne a mikroelektródán. Ennek a problémának a kiküszöbölésére nagy bemenő és kis kimenő ellenállású, emitterkövető erősítőt kell használni (lásd a sorozat 106. részét.). A mikromanipulátorok segítségével lehetséges az elektródák pontos behelyezése az agyba, ami az intracelluláris vizsgálatok esetében különösen fontos. Az erősítést követően a jelet különböző módszerekkel szűrni kell. Régebben oszcilloszkóppal és kamerával rögzítették a jeleket, manapság a jelet analóg-digitális konverterrel átalakítják, majd számítógépen tárolják.
Az egy-sejt vizsgálatok nagy térbeli és időbeli felbontással képesek információt szolgáltatni az agyműködés strukturális, funkcionális és a viselkedéssel kapcsolatos összefüggéseiről. Azzal, hogy az idegsejtek szintjén lehetséges az agyműködést vizsgálni, a kutatók képesek voltak a viselkedést és az agyterületeket egymással kapcsolatba hozni és olyan térképeket készíteni, amelyek az információnak az agyban történő áramlását követik nyomon.
Azt a sorozat 33. részéből már tudod, hogy a belső fülben a szőrsejtek tetején levő sztereocillumok elhajlanak, amikor a csiga alaphártyája megemelkedik, és az elhajlás mértékével arányos mennyiségű impulzust löknek ki magukból. Ez az impulzus sűrűség modulált elektromos jel indul el a hallóidegen keresztül az agyba. Hasonló jelenség játszódik le a látás során is. A ganglionsejtek a fotoreceptorokból származó információt sűrűség modulált elektromos impulzusokká alakítják át. Ez a jel a látóidegen keresztül jut el az agyba. Rögtön felvetődhet benned, hogy akkor a látásunk nem analóg? Erre a kérdésre ezen a ponton még egyértelműen az analóg lenne a válasz, hiszen a ganglionsejtek viselkedése a fényérzékelés alsó határa (sötétség érzet) és felső határa (vakító fény érzet) között folyamatosan a fotoreceptorok érzékelésével összhangban történik. (Az összhang nem egyenes arányosságot jelent.) A látásnak ezen a pontján még csak analógból analógba átalakításról beszélhetünk. Azt azonban már most megjegyzem, hogy a későbbiekben sem lesz majd egyszerű igen vagy nem a válasz.
Egy magyar származású amerikai tudós, Stephen William Kuffler (Kuffler Vilmos), a modern idegtudomány atyja fedezte fel 1953-ban a látás során történő információfeldolgozás legelső, és talán legfontosabb lépését.

Az imént leírt módon egy-sejt vizsgálatokat végzett, és a retinális ganglionsejtek akciós potenciáljainak másodpercenkénti sűrűségét mérte. Azt találta, hogy ezek az idegsejtek megváltoztatják tüzelési frekvenciájukat, ha a retinát kis fényfoltokkal ingerli. Ez önmagában még nem volt meglepetés, mert azt már korábban is tudni lehetett, hogy a retina reagál a fényre. A meglepetés ott kezdődött, hogy kisebb fényfoltokra inkább reagáltak ezek a sejtek, mint nagyokra, s hogy az egyes sejtek csak a retina adott helyén való ingerlésre reagáltak. Kuffler arra a következtetésre jutott, hogy azért a kis foltok hatékonyabbak, mert a ganglionsejt receptív mezője, az a kis retinális terület, amin belül egyáltalán ingerelhető, adott struktúrával rendelkezik. A receptív mező közepének ingerlésére a sejt megnöveli válaszát, míg a szélső terület ingerlésére csökkenti. Tehát a receptív mező két antagonisztikus, magyarul ellentétes működésű részből áll, s ha a fényfolt, amit ingerlésre használunk, belelóg a széli részbe, a sejt gátlás alá kerül. Ezt a jelenséget hívjuk laterális vagy széli gátlásnak.
Akkor elemezzük ezt egy kicsit részletesebben! Ehhez ismét szükség van az előző rész egyik ábrájára.
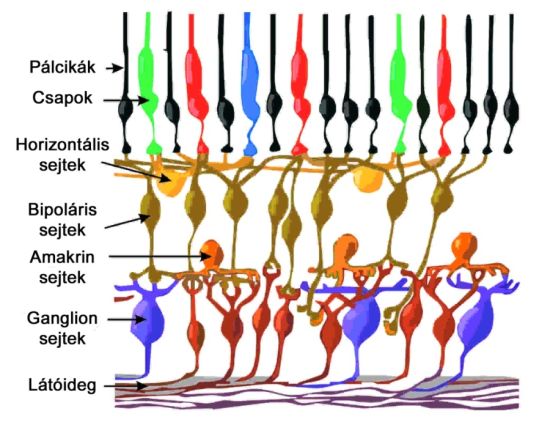
A fotoreceptorok koncentrikus felépítésű, ganglion sejtekhez kapcsolódó receptív mezőkbe rendeződnek, amelyek akár át is lapolódhatnak egymáson. A pálcikák nagyméretű, homogén mezőket alkotnak, közvetlen kapcsolatban pedig csak egyféle bipoláris sejttel állnak. Egy-egy, a pálcikákat összekapcsoló bipoláris sejthez hozzávetőlegesen 15-30 receptor tartozik. Ezek a bipoláris sejtek soha nem állnak közvetlen ingerátviteli, külföldiül szinaptikus összeköttetésben ganglion sejtekkel, a jelfolyamba minden esetben amakrin sejtek ékelődnek. A pálcikák és a hozzájuk kapcsolódó bipoláris sejtek kötegelődéséhez, egy-egy amakrin sejthez is több – nagyjából sejtenként 20 – pálcikákat összekapcsoló bipoláris kapcsolódik. Az összekapcsolódás, konvergencia a ganglion és amakrin sejtek között még ennél is nagyobb arányú lehet, esetenként egy-egy ganglion sejthez több mint száz pálcika jeleit továbbító amakrin sejt is tartozhat. A pálcikákra legjellemzőbb útvonalat követve, egyetlen ganglion sejthez akár sok tízezer receptor jele is befuthat. Ez a nagymértékű, a retinális rétegeken átívelő összekapcsolódás komoly szerepet játszik a pálcikák dominálta szkotópikus, azaz éjszakai látás jelentős érzékenységében.
A csapok alkotta receptív mezők felépítése nem homogén, hanem centrális és perifériális részből áll.
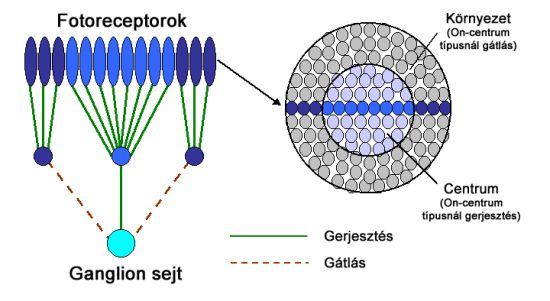
A centrális és perifériális szegmensek közötti eltéréseket a kétféle, On (be) és Off (ki) típusú bipoláris sejtek alakítják ki. Ennek megfelelően a bipoláris sejtek úgy is felfoghatók, mint egyféle előjelképző (+ és -) állomások a ganglion sejtek és csapok, vagy csapok csoportjai között. Attól függően beszélhetünk On-centrum és Off-centrum receptív mezőről, hogy a receptív mezők centrumai milyen típusú bipoláris sejten keresztül kapcsolódnak a hozzájuk tartozó ganglion sejthez.
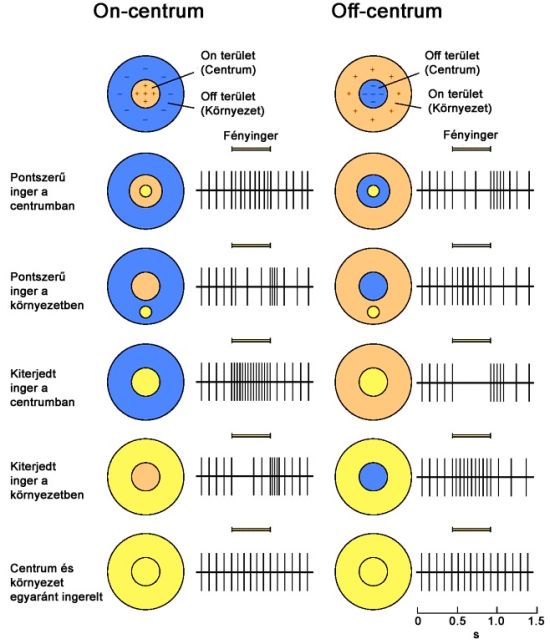
Ennek megfelelően a ganglion sejtek viselkedése az előttük lévő bipoláris sejtek viselkedését tükrözik, de egyes esetekben horizontális és amakrin sejtek is módosíthatnak a jelfolyamon. On-centrum esetben a receptor mező perifériális részének ingerlése gátolja, centrális része pedig tüzelésre – jelkibocsájtásra – készteti a ganglion sejtet. Off-centrum esetben a hatásmechanizmus éppen ellentétes, a központi terület gátló hatású, a perifériális részek pedig gerjesztik a ganglionok tüzelését.
A sárgatestben található csapok kapcsolódásai jellemzően a direkt utat követik. A receptív mezők mérete itt a legkisebb, akár egyes csapoknak is lehet külön kapcsolódási útvonala, viszont a retina perifériális részei felé haladva a receptorok egyre nagyobb méretű receptív mezőket alkotnak. Ez az egyik oka annak, hogy perifériális látásunk térbeli felbontása jóval gyengébb, mint azt a látóterünk közepén tapasztaljuk. Ahogy a pálcikák alkotta mezők esetében, úgy a nagyobb méretű csap receptív mezők kialakításában az egyes receptorokból érkező jelek összefűzésével, esetenként a jelfolyam módosításával az amakrin és horizontális sejtek is szerepet játszanak.
Ha eddig tudtad követni, igazán elismerésem, pedig most jön a java!
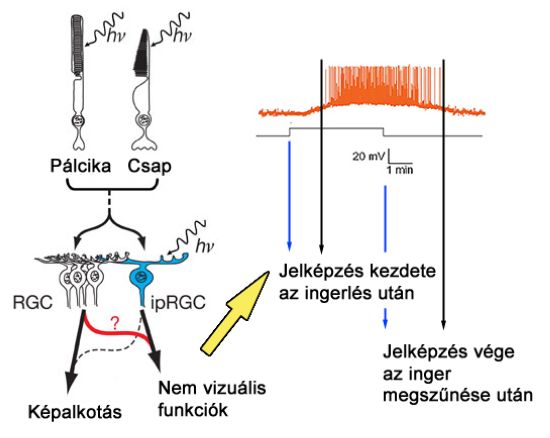
A ganglion sejtek alkotják a látókéreg előtti utolsó állomást, így ezen sejtek akciós potenciáljai képezik a retina kimeneti jelét. Ganglion sejtekből négyféle típust különböztetünk meg: PC (vagy P – parvocelluláris), MC (vagy M – magnocelluláris), KC (vagy K – koniocelluláris) és ipRGC (Photosensitive, azaz fényérzékeny) típust. Az PC és MC ganglion sejteknek centrális és perifériális részből álló receptív mezői vannak. Az előbbiekhez kisebb méretű, az utóbbiakhoz nagyobb kiterjedésű receptív mezők tartoznak, és amíg a PC ganglion sejtek színlátásunk alapjait képezik, és csak nagyon csekély mértékben érzékenyek a kontrasztváltozásokra, az MC típusúak nem játszanak fontos szerepet a színérzékelésben. A KC típusú sejtek a többi ganglion típushoz képest kisméretűek, receptív mezejük csak centrális részt tartalmaz, amely a kék csapokhoz kapcsolódva mindig on-, vörös vagy zöld csaphoz csatlakozva pedig mindig off-típusú. Szerepük teljes mértékben még nem tisztázott, de a kontrasztérzékelésben van funkciójuk. Az ipRGC típusú ganglion sejtek receptív mezeje sokkal heterogénebb, csapokat és pálcikákat egyaránt tartalmaz, és az általuk továbbított ingerületek nem a vizuális ingerek kialakításában játszanak szerepet, hanem a fény egyéb hatásainak formálásáért felelnek.
A kép világosságjele, tehát az a jel, ami csak a szürke árnyalatait – a fehértől a feketéig – tartalmazza, akromatikus csatornajel néven szerepel a tudományban. Ez a környezet objektumainak világosság jellemzőivel kapcsolatban kódol információt. Az emberi szem világosságérzékelésének spektrális vizsgálatai során kimutatták, hogy a nappali látásérzékelésünk hullámhosszfüggő hatékonyságát leíró függvény jól közelíthető a vörös-érzékeny (L) és zöld-érzékeny (M) csapok érzékenységi karakterisztikáinak súlyozott összegével. Ez az összegzés az erre specializálódott ganglion sejteken keresztül történik. A világosságjel kialakításában szerepet játszó ganglion sejtek receptív mezejének mind külső, mind centrális részén L és M csapok is megtalálhatók. Ezek jellemzően nagy kiterjedésű, széles kapcsolatrendszerű mezők, amelyek közt mind On-centrum, mind Off-centrum típusúak is megtalálhatók. On-centrum esetében a receptív mező középpontjának ingerlésére nő meg a ganglion sejtek tüzelési frekvenciája, így ez a mechanizmus a sötét háttér előtt megjelenő világos objektumok érzékelését végzi. Off-centrum esetben a hatás pont az előző fordítottja, a környezetet alkotó csapok ingerlése gerjesztő, míg a centrumra eső fény gátló hatású a ganglion sejt kimenetére nézve, így a világos háttér előtt megjelenő sötét objektumok idéznek elő magasabb tüzelési frekvenciát. Különbség még az On-centrum és Off-centrum mezők között, hogy működésük jellegéből adódóan utóbbiak kontrasztérzékenysége nagyobb. Egyenletesen világos környezet esetében mindkét mezőkialakítás kimenete átlagos frekvenciával tüzel, hiszen a gerjesztő és gátló mezők egyszerre ingereltek, egyenletesen sötét háttér esetén pedig egyik esetben sincs jelképzés. Az akromatikus csatornajelet kialakító mechanizmus hatására jön létre a laterális gátlásnak nevezett folyamat, amelynek segítségével a receptor mezők szerkezetéből adódó következmények jól szemléltethetőek. Az ilyen és ehhez hasonló retinális feldolgozási folyamatok esetenként optikai csalódásokon keresztül érhetőek tetten.
Ez a sok rizsa ahhoz kellett, hogy visszajussunk a laterális gátlásig. Nézzed meg egy híres vizuális illúzió kapcsán, hogy mit is tud a laterális gátlás!
Ludimar Hermann 1870-ben fedezte föl a róla elnevezett vizuális illúziót.

A Hermann-rácsot nemcsak azért lehet szeretni, mert vicces és játékos, hanem azért is, mert remekül feltárja a világosság észlelés mechanizmusait. Vagy legalábbis ezek egy részét.

A fenti ábrát vizsgálva feltűnhet, hogy a látómezőnk perifériális részén a csomópontokban sötét foltokat érzékelsz, holott tudod, hogy a világos sávok kialakítása homogén. Azt is észreveheted, hogy a látómezőnk centrális részén a jelenség nem figyelhető meg, tehát ha koncentráltan nézel egy kereszteződést, az nem szürkül el. A foltok kialakulásának magyarázata az, hogy amikor a szomszédos sötét mezők oldalai közti világos sávok képe On-centrum típusú receptív mezőre esik a retinán, a mezőhöz kapcsolódó ganglion sejt erős gerjesztést kap, hiszen a gátló területekre csak vertikális vagy horizontális irányban esik fény, így a gátló mező területének nagy része nem kap gerjesztést. A csomópontokban, azaz a sötét mezők csúcsainál viszont a perifériális gátló mezőterület ingerlése az itt horizontális és vertikális irányban is jelenlévő csíkozat miatt kétszeres az élek mentén kialakuló körülményekhez képest, így az agy azt az információt kapja, hogy a csúcsok közti terület sötétebb. Ez kontrasztkiemelő hatást eredményez, melynek következtében a kiterjedtebb sötét határral rendelkező világos képrészek intenzívebbnek tűnnek. Az akromatikus csatorna jeltovábbítása vastagabb, magnocelluláris idegpályákon, MC típusú ganglion sejtek közreműködésével történik, így a jelfolyam gyorsabb, de jóval kisebb térbeli felbontású, azaz nem képes a vizuális ingerek finom részleteinek érzékelésére. A retina centrális részére nem jellemző ez a fajta mezőszerkezet, hiszen a látómezőnk közepére pozícionált csomópont esetében nem figyelhető meg a periférián jól érzékelhető gátló hatás. Ennek oka, hogy a retina centrális részén a receptív mezők finomabb szerkezetűek. Ez a felfogás azért is nagyon vonzó, mert közvetlen kapcsolatot teremt a fiziológiai jelenségek világa és az élmény világa között. A Hermann-rács illúziójának magyarázata tehát, úgy tűnik, retinális szinten van.
A későbbi kutatások kimutatták, hogy a laterális gátlás nem elégséges, mivel az illúzió még akkor is fennmarad, amikor azonos világosságú területek találkoznak, vagy a rács geometriája megváltozik. Az elsődleges látókéregről (V1) lesz még szó. Ezen kéreg idegsejtjeinek orientációs érzékenysége is hozzájárul az illúzióhoz. Ezek a sejtek nemcsak világosságkülönbségeket dolgoznak fel, hanem irányérzékenyek, így a rács metszéspontjainál eltérő mintázatot észlelhetnek, mint a rácsvonalak mentén. A vizuális rendszer hajlamos „kitölteni” az észlelési hiányokat. A Hermann-rács illúzió során az agy prediktív, jósló mechanizmusai feltételezik, hogy a metszéspontok sötétebbek, mert ott nagyobb kontrasztra számítanak. A prediktív kódolás ismerős lehet számodra pl. a hangtechnikában használatos .flac formátumból (lásd a sorozat 96. részét). Amikor fókuszálunk egy pontra, a figyelem segít elnyomni az illúziót, mert részletesebb feldolgozás történik a fókuszált területen.
A modern, számítógépes neurális hálózati modellek, amelyek elsősorban a tanulást modellezik, képesek reprodukálni a Hermann-rács illúziót, ami sejteti, hogy tanulásalapú vizuális feldolgozás is képes generálni ilyen illúziókat. Ez alátámasztja, hogy az illúzió nemcsak a szem strukturális sajátosságaiból ered, hanem a komplex, tanult agyi feldolgozás terméke.
Most egy színes képet mutatok neked, amelyen szintén működik az illúzió. Ez a kép azért érdekes, mert az én kérésemre gyártotta a ChatGPT. A sorozat ezen része elkészítéséhez egyébként is használtam mesterséges intelligenciát, de minden állítást ellenőriztem.

A ChatGPT felajánlotta, hogy animációkat készít nekem, válasszak, hogy melyik fajtát. Én az összeset kértem tőle. Először azt írta, hogy elküldi az egyes változatok kép-összetevőit, és én egy animáció szerkesztővel rakjam össze. Noha van ilyen szerkesztő progim, üzentem, hogy rakja össze ő! És láss csodát, megcsinálta! Nem mindegyik lett meggyőző, de négy igazán cseles. Az első animáció azt mutatja be, hogy ha a négyzetek lassan eltolódnak, a sötét foltok mozgása látványosabb lesz.

A második először fekete-fehér, majd színes és inverz változatokat mutat animált ciklusban.

A harmadikban a rács elmosódásának változtatásával láthatod, mikor szűnik meg az illúzió.

A negyedikben pedig egy „szem” mozgatásával derül ki, hogyan tűnnek el a foltok, ha ráfókuszálsz.

Érdekes átalakítást hajtott végre a Hermann-rácson Geier János matematikus 2004-ben.

A rácsminta egyeneseinek hullámossá tételével eltüntette az illúziót. Ez azért különösen érdekes, mert bár az eddig bemutatott retinális receptív mező modell szerint a kereszteződésekben foltoknak kellene megjelenniük, ebben a speciális esetben még sincs így, a foltok megszűnnek. A laterális gátlásról és a retinális receptív mezőkről mondottakat nem cáfolja az új illusztráció, sőt ezek alapján lehetséges az illúzió értelmezése. Ez is azt bizonyítja, hogy a Hermann-rács nem pusztán a retinális feldolgozás eredménye, hanem agykérgi folyamatok is szerepet játszhatnak benne.
A laterális gátlás jelentősége természetesen sokkal nagyobb, mint pusztán a vizuális illúziók területén való alkalmazásuk. A laterális gátlás hasonló elven alapul, mint amikor egy hangmérnök kiemeli a szólista hangját a vele együtt éneklő, nagylétszámú kórusból, mégpedig úgy, hogy a kórusmikrofonok jelét visszaveszi. Nem fordítva, nem a szólista mikrofonjának jelét erősíti, mert az torzításhoz vezetne. Az idegrendszer a fontos, időben vagy térben változó ingerek kódolására hangolódott. A változások kiemelésének egyik legfontosabb eszköze a laterális gátlás. A látás esetén ez úgy valósul meg, hogy az idegsejt (például a retinális ganglionsejt) csak akkor változtat aktivitásán, ha a receptív mezőn belül változás van, például hirtelen változik a fény intenzitása. Ha nincs változás, akkor az aktivitás sem változik – például, amikor a receptív mező egy egyszínű alakzat belsejébe esik. A következő, az egyszerűség kedvéért fekete-fehér ábrán azt láthatod, hogy a retinális ganglionsejtek a laterális gátlás egyszerű mechanizmusát felhasználva, mit hoznak létre a retinára vetülő képből.

Ez bizony igen erős egyszerűsítés, nagyon veszteséges tömörítés. Márpedig a hangtechnikában nagyon rossz néven vettük az ennél sokkal kisebb tömörítést is. Csakhogy az agyunk részben a genetikai kódolás segítségével tudja, az egyedfejlődés során pedig megtanulja, hogy mit kell majd láttatnia e tömörítés feldolgozása során. Miért is fontos ez a tömörítés, miért nem küldi a szem a kép pontos és hű, pontról pontra megegyező másolatát az agynak? Azért, mert akkor a szemenként 100 millió receptor 100 millió képpontját egymástól függetlenül kellene átvinni az agy felé. Ehhez nagy „sávszélesség” kellene, a látóideg olyan vastag lenne, és akkora koponya kellene a befogadásához, hogy az a szülőcsatorna jelentős megnövekedését igényelné, ami viszont a két lábon járás képességének meglehetős akadálya lenne. Csakhogy, ha soha nem használja a látáshoz az idegrendszerünk mind a százmilliót, akkor minek van ennyi belőle? Nos, a természet igen bőkezű, óriási szerkezeti redundanciával (lásd a sorozat 95. részét) ajándékoz meg minket, s ez nem csupán a szaporodásra érvényes, amikor sokmillió gecikéből csak egy igazán fürge termékenyítheti meg a petesejtet.
A retina ideghálózata, nagyrészt a ganglionsejtek receptív mező struktúrája következtében, elvégzi a retinakép feldolgozásának első jelentős mozzanatát, a lényeges változások kiemelését. Ezek a változások általában hirtelen kontrasztugrások, melyek ott jelentkeznek, ahol egy tárgyhoz vagy élőlényhez tartozó felszín véget ér vagy takarásba kerül egy másik tárgy által. Tehát fizikai világunk egységeinek, a tárgyak és előlények határoló felszíneit, vonalait, kontúrjait küldi tovább a retina az agynak. Ez azonban még csak az „első közelítés”, mert a ganglionsejtek receptív mező ablakai igen kicsik, és a szomszédos ablakok információja csak az agykéregben kombinálódik. A retina vázlata a tárgyakról még nagyon durva és zajos, de az agykéreg tudja, mit kezdjen majd vele, hogyan válogassa ki a zajból a valódi információt, hogyan egyeztesse a kontúr, a szín, a mélység és a mozgás információját, és hogyan rakja össze az elemi információk alapján a számodra fontos valóság egy darabját.
De azért van itt még egy kérdés. Ha az agynak elég egy ennyire tömörített adatsor, akkor, ha valamiyen módon az autó tömörített képét tudnánk a sárgatestre vetíteni, a szervezetünk előállítaná az autó teljes, részletes képét? Noha erre vonatkozóan nem találtam kísérleti eredményeket, de valószínűleg nem.
Viszont ne becsüld le az agyad képességeit! Gyakorta egy-egy egyszerűsített vázlatból, kevésbé fejlett ábrázolásokból, amelyek lényeges képi információkat tartalmaznak, fel tudjuk ismerni, hogy mit is akart mondani az alkotó, hiszen ismerjük a kép közlendőjéhez tartozó jelentésbeli kódot.

Ez még akkor is igaz, ha a képen szándékos torzítások vannak.

Sőt. Az alkalmazott mateknak van egy ága, az ábrázoló geometria. Ennek lényege, hogy az ábrázolandó tárgyról bizonyos szabályok segítségével lehet készíteni két vagy három síkbeli képet. Egyet felülnézetből, egyet oldalnézetből egyet pedig szemből. Ha egy gömböt a forgástengelyére merőleges síkkal metszünk el, minden nézete kör lesz.
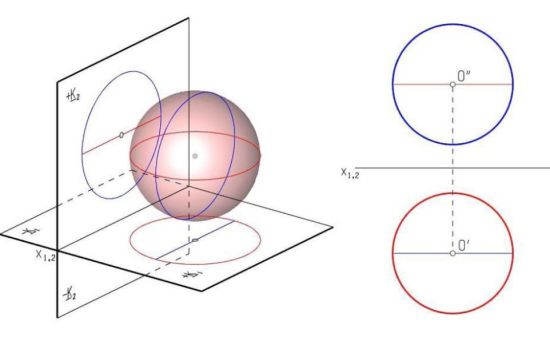
Az emberek egy része a nézeti képeket megtekintve, látja a gömböt, s ennél bonyolultabb formákat is. Nemcsak tudja, hanem tényleg látja, a képzeletében megjelenik a tárgy. Bevallom, én soha nem láttam, ezért bemagoltam az egész Műszaki rajz tankönyvet, s harmadszorra meg is kaptam a vizsgán az elégségest, a tanárom megjegyzésével, hogy csak azért engedett át, mert ennyire fantáziátlan, szelektív vakkal még sosem találkozott.