
Cs. Kádár Péter - XXI. századi Diszkónika, 499. Látópályára fel!
A retinát a látókéreggel a látópálya köti össze.
A felépítésének oka részben ismeretlen, és a szerkezet érthető ismertetése sem egyszerű. Nem árt, ha a sorozat néhány előző részét ismét elolvasod.
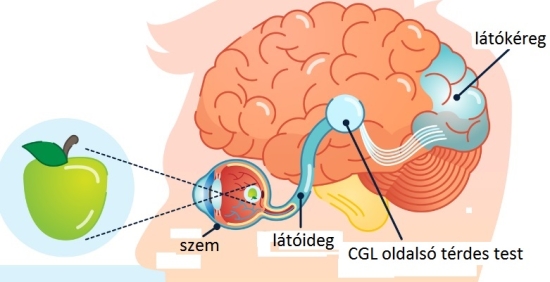
A látóideg az ideghártya nagy ganglionsejtjeinek axonjaiból szedődik össze a látóidegfőnél. Ez a szem hátsó pólusától 3-4 milliméterre befelé található. A vizuális rendszer a legkomplexebb neurális rendszer. A látóideget kb. egymillió rost alkotja. Az alábbi animáció a fényinger útját követi a látóidegig.
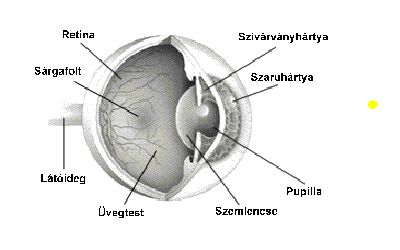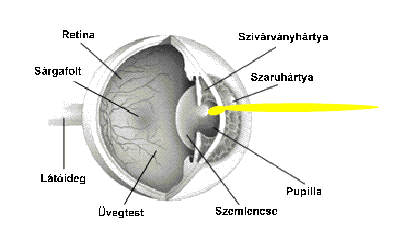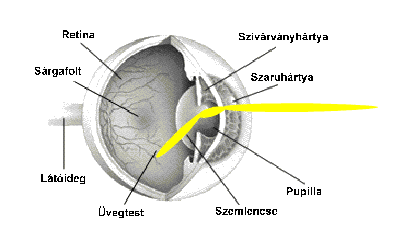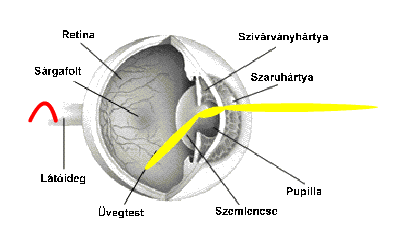
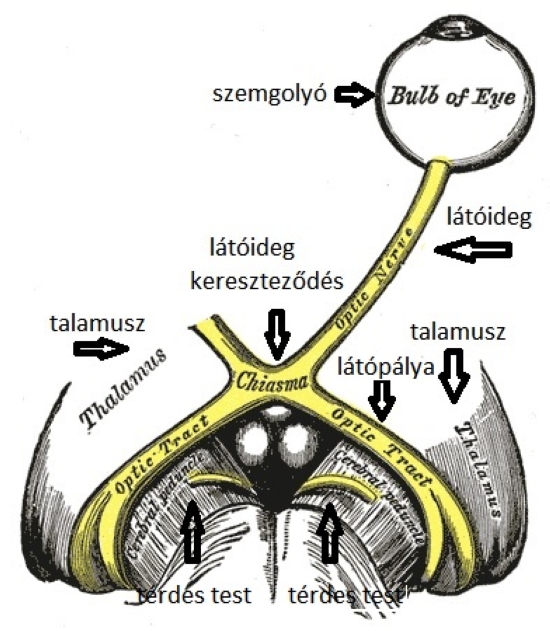
A szemen belül a ganglionsejtek axonjait, nyúlványait nem borítja mielin (velőhüvely, amely szigeteli az axont, növeli a vezetőképességet). A látóidegek a látóideg-kereszteződésben futnak össze, ahol az idegrostok átrendeződnek. Mégpedig úgy, hogy egyesek mindig az azonos oldalon maradnak, míg mások átkereszteződnek. A rostok egy része nem kereszteződik (ipszilaterális rostok; az „ipsi” jelentése „ugyanaz”). Az ipszilaterális rostok mindig a halántékhoz közeli retinafélből erednek, mégpedig mindkét szem esetében. Az ellenkező oldalra futó vagy keresztezett rostok (kontralaterális rostok; a „kontra” jelentése „ellen-, szemben”) az orr felé eső retinafélből erednek. Az ember két szeme axonjainak mintegy fele átkereszteződik; a kereszteződő, illetve változatlanul tovahaladó rostok a retinának a sárgatestre merőleges tengelyétől a halánték, illetve az orr felé eltérőek. A látóideg-kereszteződésben a kereszteződő és a nem kereszteződő rostok kapcsolódnak, majd felfelé, az agy felé haladnak tovább. A látóideg-kereszteződés felett az axonkötegeket látópályáknak vagy látókötegeknek (tractus opticus) nevezik.
A fenti képen ábrázolt talamusz az agy mélyén ülő sejtcsoport, ami az agykéreg felől kiinduló és az oda tartó idegi jelek “átkapcsoló állomásaként” funkcionál, a rajta keresztül haladó idegpályák integrálását végzi.
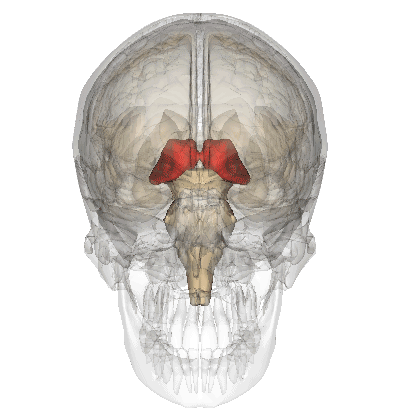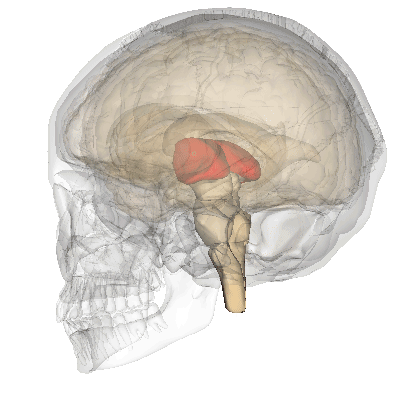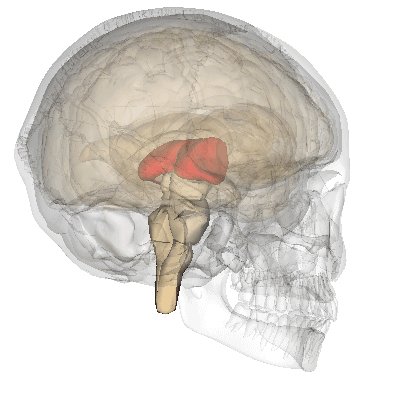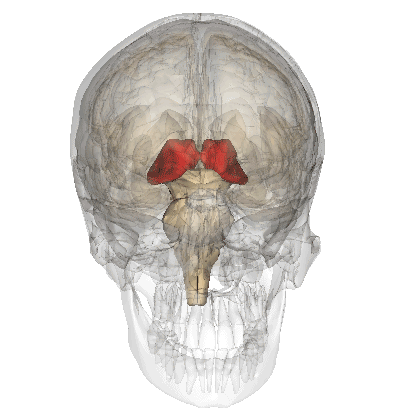
A szaglás kivételével az összes érzékszerv információja beérkezik a talamuszba, amelyek innen továbbítódnak a többi agyterület felé.
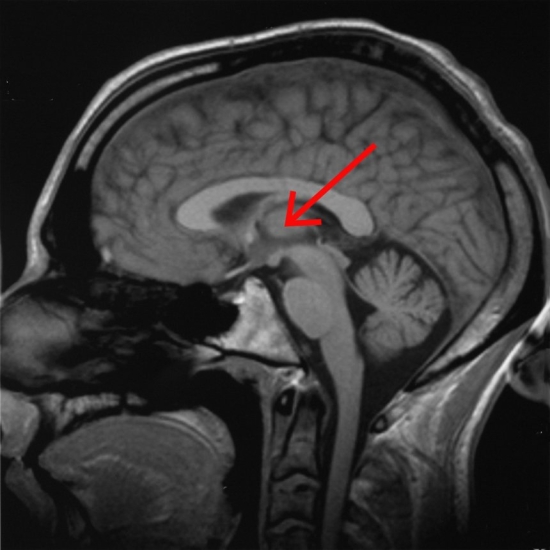
Ehhez hasonlóan, ám ellentétes irányban vesz részt a mozgás vezérlésében is. A talamusz tehát olyan, mint egy hatalmas, kusza kereszteződés, ahol sok idegi információ összegződik és kapcsolódik át a központi idegrendszer többi része felé; mintha az információk itt rendeződnének a nekik megfelelő sávokba, ahonnan úti céljuk felé kanyarodhatnak.
A látóideg rostjainak jelentős része az oldalsó térdes test (anatómiai nevén corpus genigulatum laterale – CGL vagy Lateral Geniculate Nucleus – LGN) felé halad, kisebb hányada a középagy tetején levő felső dombba, a colliculus superiorba (CS) vezet. A CGL és CS területeire a retina axonjai „rávetítik” a retinális információt, ezeket a helyeket projekciós területeknek is nevezik a látással foglalkozó pszichológusok. Ennek a két vizuális központnak gazdag összeköttetés rendszere van, és a két központ összehangoltan működik. A középagy struktúráinak fontos szerepük van a pupillaméret szabályozásában, a saját mozgás érzékelésében, valamint a látás és a testtartás, a testmozgás összehangolásában. Vizuális-poszturális alkalmazkodásnak hívják, akik mindenképpen dicsekedni akarnak a latin tudásukkal.
A CS sejtjeinek receptív mezői viszonylag rosszul meghatározottak, ez kifejezetten érvényes az OFF és az ON sejtekre. A CS részt vesz a szemmozgások vezérlésében, a szemmozgások indításában, a mozgások irányának és terjedelmének vezérlésében.
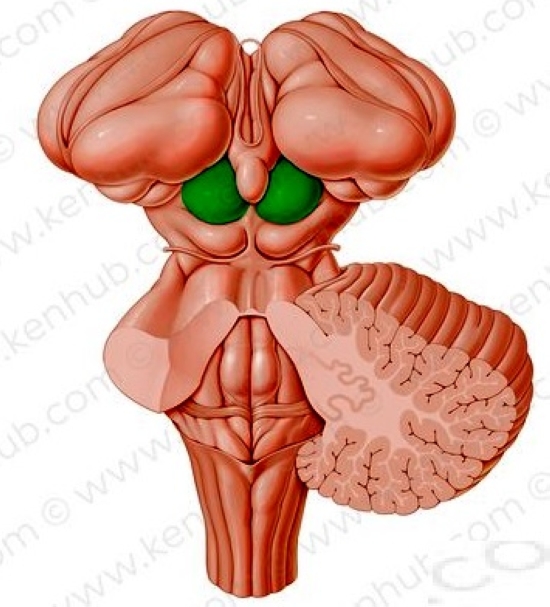
A CS a hallási és vizuális érzékletek közötti integrációban is meghatározó szerepet tölt be. A CS-ben olyan multiszenzoros sejtek találhatók, amelyeknek van vizuális és hallási bemenetük. A multiszenzoros sejtek választevékenysége leginkább az azonos forrásból származó, tehát a tér azonos területéről érkező hangokra és látási ingerekre terjed ki. A CS legfontosabb feladatai közé tartozik a legélesebb, fixációs ponttól távolabbi tárgyak detektálása, az ezekre irányuló szemmozgások irányítása.
A CGL-nek sajátos rétegszerkezete van: hat, a közepénél hajlított, egymás felett elhelyezkedő rétegből áll. Ennek a hajlított, térdszerű formának köszönheti nevét is: térdes test. A CGL első két rétegét nagyobb sejtek alkotják, mint a felette elhelyezkedő négy rétegét. Ezeket a nagyobb sejtekből álló rétegeket nevezzük magnocellulárisnak (M), a kisebb sejtekből állókat pedig parvocellulárisnak (P). A P retinális ganglion- sejtek adják a CGL parvocelluláris, az M sejtek pedig a CGL magnocelluláris rétegeinek a bemenetét. A CGL páros struktúra, mindkét szem küld ezekhez információt; az azonos oldalit az azonos oldali CGL-nek, az ellenoldalit a másik oldalon lévő CGL-nek. A CGL a látópályának olyan utolsó átkapcsolódása, ahol a befutó információk egyfajta analóg válogatása történik.
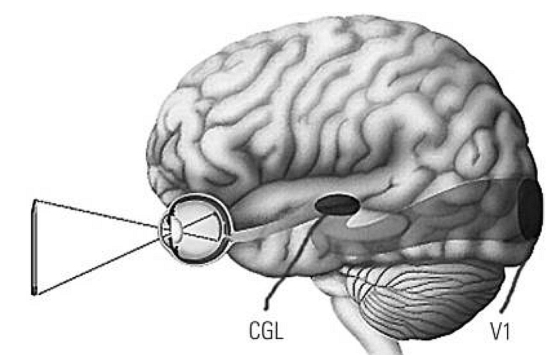
A befutó információk szabályos elrendezése a CGL-ben a vizuális feldolgozásnak azokat a további fázisait segíti, amelyek az elsődleges látókéregben, a V1-ben zajlanak le. A fenti kép ezt foglalja össze. A tárgyról a szem optikája képet formál, mely a szem hátsó falán lévő retinára vetül. A látóideg, mely a retinális ganglionsejtek kivezető huzaljait (axonok) tartalmazza, egy agykéreg előtti idegmagba juttatja a kivonatolt információt. Ebből a magból (CGL) azután egy átkapcsolással az agykéreg elsődleges látókérgébe (V1) vetül a látópálya.
Megértelek, ha tucatnyi új fogalom, elnevezés után szomorúan nézel magad elé, és például nem érted, miért van többszöri átkapcsolás. Ez a kérdés már a hallópálya ismertetésekor (lásd a sorozat 34-35. részét) is fölmerülhetett, és esküszöm, hogy több szakembertől megkérdeztem, de nem kaptam rá választ. Pedig mindennek van oka, csak úgy tűnik, ezekkel a kérdésekkel kevéssé foglakoznak.
Természetesen voltak kezdeményezők, akik például a sorozat 497. részében említett Stephen Kuffler nyomdokain haladtak tovább. David Huntel Hubel és Torsten Wiesel, akik az 1950-es években még igen fiatal kutatók voltak, nekiálltak a Kuffler-féle méréseket most már nem a retinán, hanem az agykéregben elvégezni.

Nagy merészség volt ez, mert akkoriban csak annyit lehetett tudni, hogy az agy tarkólebenyében van egy látással foglalkozó terület.
Azt viszont, hogy e terület idegsejtjeinek pontosan mi is a dolguk, senki nem tudta. Azt pedig elég merész elképzelés volt, hogy külső vizuális ingerlésre ezek a sejtek reagálni fognak. Hubel és Wiesel szorgalmasan alakították, változtatták az ingeradás feltételeit. Macskák, később majmok agyát vizsgálták. Az akkor divatos diaképekhez fémkeretes üveglapokat használtak, s arra különböző méretű foltokat ragasztottak fel.
Egyszer csak találtak egy olyan sejtet, kísérleteikben a 3006-os számút, amely mintha válaszolt volna az egyik diakeretben elhelyezett foltra. Mintegy kilenc órán át ugyanazzal a sejttel próbálkozva rájöttek, hogy a sejt nem is a foltra, hanem a diakeretre válaszol, és annak is csak a vízszintes irányú élére. 1981-ben Nobel-díjat kaptak, mert megfogalmazták a modern idegtudomány egyik legfontosabb működési törvényszerűségét, a neuronális szelektivitás elvét. Ez azt jelenti, hogy az agykéreg idegsejtjeinek egy része a fizikai ingerek egyes tulajdonságaira hangolódik, s ezekre szelektíven reagál. A kísérletben szereplő 3006-os sejt a tarkólebenyben adott irányú éldarabokra reagál, és leginkább csak azokra, tehát orientáció-, azaz irányulásszelektív. A két tudós arra a felismerésre jutott, hogy a vizuális információfeldolgozás az agykéregben a retina körkörös receptív mezőit követi. A feldolgozás egyre bonyolultabb szintjei hierarchikusan követik egymást. Tény, hogy a Hubel és Wiesel által leírt, a vonal irányulására (orientációjára) szelektív neuronok valóban hierarchikusan is bonyolultabbak, mint retinális elődjeik, s hogy a vizuális feldolgozórendszer magasabb szintű agykérgi területein, távolabb a tarkólebenytől, ennél még bonyolultabb működésű sejtek vannak.